
产品中心
技术交流

扫描二维码
抗原交叉呈递:外源抗原如何“走后门”激活 CD8⁺ T 细胞?
文章来源公众号:lmmunoUpdate 作者:lmmunoUpdate
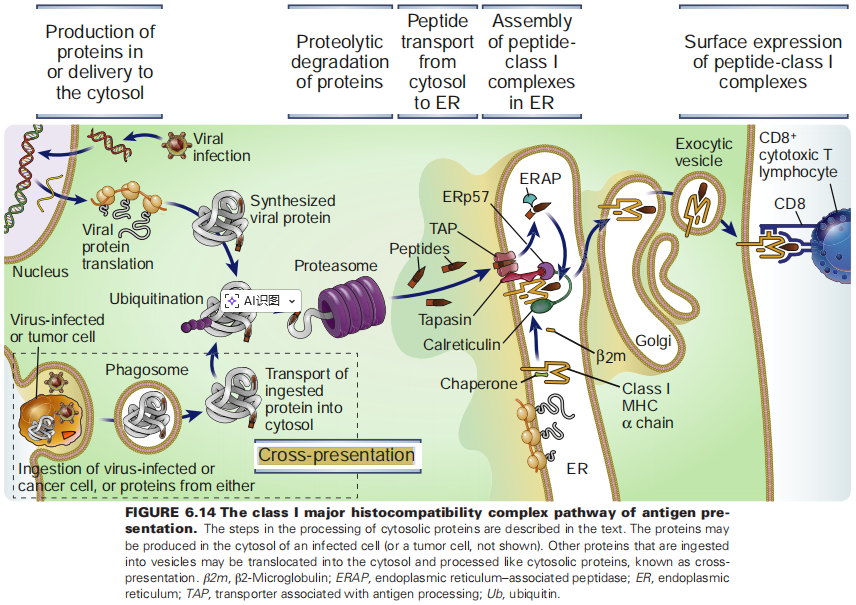
(图片来源:Cellular and Molecular Immunology)
在正常情况下,抗原呈递细胞(APC,如树突状细胞)遵循两条严格分开的流水线来处理抗原,就像机场的国内和国际安检通道一样分开:
MHC-I 途径(处理“内鬼”):
MHC-II 途径(处理“外敌”):
想象一下这种情况:你感染了一种病毒(比如乙肝病毒或流感病毒),这种病毒只感染你的上皮细胞或肝细胞,但不感染专业的抗原呈递细胞(比如树突状细胞,DC)。
这就是需要“走后门”的时候了。 免疫系统必须有一种机制,让DC能够获取外部的病毒抗原(比如吃掉一个死掉的被感染细胞),但却能把这些抗原呈递给 CD8⁺ T细胞。
这就是交叉呈递(cross-presentation)[1–3]。
真实世界里,并不是所有树突状细胞(dendritic cell,DC)都同样擅长交叉呈递。大量小鼠和人类研究表明,经典 1 型树突状细胞(conventional type 1 DC,cDC1,典型表型为 XCR1⁺、CLEC9A⁺)是体内最专业的交叉呈递细胞,在抗病毒感染和抗肿瘤免疫中尤其关键。[2,4–6]
其他 DC 亚群(如 cDC2)以及部分组织巨噬细胞,在特定炎症环境或组织部位也能进行交叉呈递,但效率和条件依赖性往往不如 cDC1,更多像是“兼职保安”。[2,7]
从“技术路线”上看,外源抗原要想挤进 MHC I,有两条主要通道:
细胞质途径(cytosolic pathway)——绕一圈再回来
液泡/囊泡途径(vacuolar pathway)——“就地消化、原地装配”
要保证“走后门”成功,还有一个关键“暗箱”:不能把抗原一锅炖烂。cDC1 利用 NADPH 氧化酶 NOX2 往吞噬体里释放少量活性氧,把囊泡 pH 稍微“碱性化”,减慢蛋白降解速度,既保留足够抗原,又能生成适合 MHC I 的肽段。[4]
那为什么免疫系统要费这么大劲开这道“后门”?
用一句更生活化的话收束:交叉呈递就是免疫系统给外源抗原开的一条 VIP 后门,把本来只属于“内部员工”的 MHC I–CD8⁺ T 细胞通路,扩展成了对病毒、肿瘤和某些疫苗抗原的“外来访客专线”。理解这道“后门”怎么开、由谁控制,也就抓住了抗肿瘤免疫和下一代疫苗设计中的关键杠杆。
 粤公网安备 44010602000473号
粤ICP备09063491号
营业执照
粤公网安备 44010602000473号
粤ICP备09063491号
营业执照

